The human genome encodes hundreds of proteins that ferry ions across membranes. How did they get here? How do they work?
As macroscopic beings, we generally think we are composed of tissues like bones, skin, hair, organs. But this modest apparent complexity sits atop a much greater and deeper molecular diversity- of molecules encoded from our genes, and of the chemistry of life. Management of cellular biochemistry requires strict and dynamic control of all its constituents- the many ions and myriad organic molecules that we rely on for energy, defense, and growth. One avenue is careful control across the cellular membrane, setting up persistent differences between inside and outside that define the living cell- one may say life itself. Typical cells have higher levels of potassium inside, and higher levels of sodium and chloride outside, for example. Calcium, for another example, is used commonly for signaling, and is kept at low concentrations in the cytoplasm, while being concentrated in some organelles (such as the sarcoplasmic reticulum in muscle cells) and outside.
All this is done by a fleet of ion channels, pumps, and other solute carriers, encoded in the genome. We have genes for about 1,555 molecule transporters. Out of a genome of about 20,000 genes, this represents a huge concentration(!) of resources. One family alone, the solute carrier (SLC) family, has 440 members. Many of these are passive channels, which just let their selected cargo through. But many are also co-transporters, which harness the transport of one ion with that of another which may have an actively pumped gradient across the membrane and thus provide an indirect energy source for transfer of the first ion. The SLC family includes channels for glucose, amino acids, neurotransmitters, chloride, cotransport (or anti-transport) of sodium with glucose, calcium, neurotransmitters, hydrogen, and phosphate. Also, metals like zinc, iron, copper, magnesium, molybdate, nucleotides, steroids, drugs/toxins, cholesterol, bile, folate, fatty acids, peptides, sulfate, carbonate, and many others.
It is clear that these proteins did not just appear out of nowhere. The "intelligent" design people recognize that much, that complex structures, which these are, must have some antecedent process of origination- some explanation, in short. Biologists call the SLC proteins a family because they share clear sequence similarity, which derives, by evolutionary theory, and by the observed diversification of genes and the organisms encoding them over time, from duplication and diversification. This, sadly, is where the "intelligent" design proponents part ways in logic, maintaining perhaps the most pathetic (and pedantic) bit of hooey ever devised by the dogmatic believer: "specified information", which apparently forbids the replication of information.
However, information replicates all the time, thanks to copious inputs of energy from the sun, and the advent of life, which can transform energy into profusions of reproduced/replicated organisms, including replication of all their constituent parts. For our purposes, one side effect of all this replication is error, which can cause unintended replication/duplication of individual genes, which can then diverge in function to provide the species with new vistas of, in this case, ionic regulation. In yeast cells, there are maybe a hundred SLC genes, and fewer in bacteria. So it is apparent that the road to where we are has been a very long one, taking billions of years. Gene duplication is a rare event, and each new birth a painful, experimental project. But a family with so many members shows the fecundity of life, and the critical (that is, naturally selected) importance of these transporters in their diverse roles throughout the body.
 |
| A few of the relatives in the SLC26A family, given in one-letter protein sequence from small sections of the much larger protein, around the core ion binding site. You can see that they are, in this alignment, very similar, clearly being in the same family. You can also see that SLC26A9 has "V" in a position in alpha helix 10, which in all other members is a quite basic amino acid like lysine ("K") or arginine ("R"). The authors argue that this difference is one key to the functional differences between it and SLC26A6. |
A recent paper showed structures for two SLC family members, which each transport chloride ion, but differ in that one exchanges chloride for bicarbonate, while the other allows chloride through without a matched exchange (though see here). SLC26A9 is expressed in the gut and lung, and apparently helps manage fluid levels by allowing chloride permeability. It is of interest to those with cystic fibrosis, because the gene responsible for that disorder, CFTR, is another transporter, (of the ABC family), and plays a major role doing a similar thing in the same places- exchanging chloride and bicarbonate, which helps manage the pH and fluidity of our mucus in the lung and other organs. SLC26A9, having a related role and location, might be able to fill some of the gap if drugs could be found to increase its expression or activity.
SLC26A6 is expressed in the kidney, pancreas, and gut, and in addition to exchanging bicarbonate for chloride, can also exchange oxalate, which prevents kidney stones. Very little, really, is known about how all these ion transporters are expressed and regulated, what differentiates them, how they relate to each other, and what prompted their divergence through evolution. We are really just in the identification and gross characterization stage. The new paper focuses on the structural mechanisms that differentiate these two particular SLC family members.
 |
| Structure of two SLC transporters, each dimeric, and superimposed. The upper parts are set in the membrane, with the lower parts in the cytoplasm. The upper parts combine two domains for each monomer, the "core" and "gate" domains. The channel for the anion threads within the center of each upper part, between these two domains. Note how structurally similar the two family members are, one in green+gray, the other in red+blue. |
 |
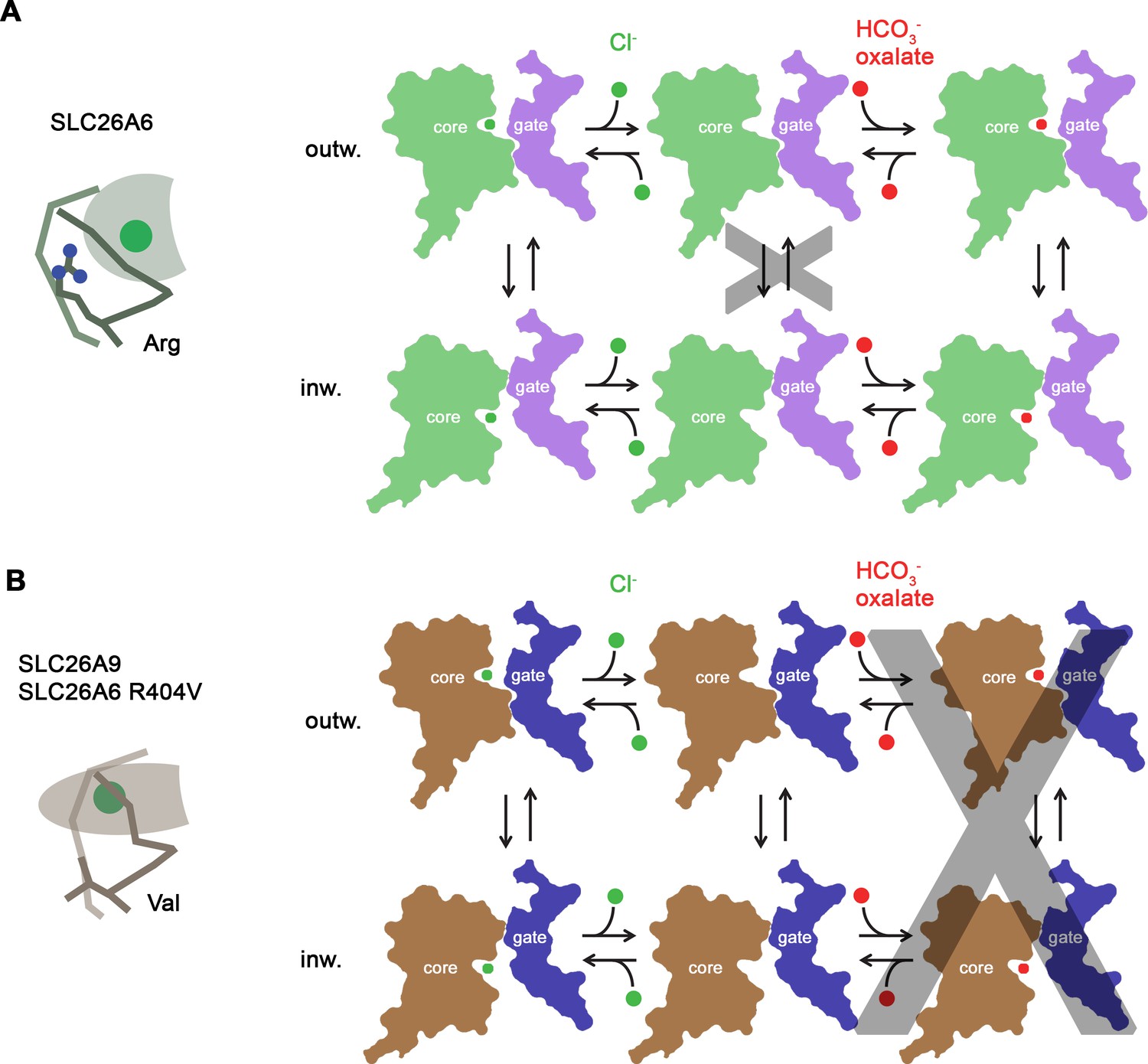
| Schemes of how SLC26A6 works. The gate domain (purple) is stable, while the core domain (green) rocks to provide access from the ion binding site to either outside or inside the cell. |
Like any proper ion channel, SLC26A6 sits in the membrane and provides a place for its ion to transiently bind (for careful selection of the right kind of ion) and then to go through. There is a central binding site that is lined specially with a few semi-positively charged amino acids like asparagine (N), glutamine (Q) and arginine (R), which provide an attactive electronic environment for anions like Cl-. The authors describe a probable mechanism of action, (above), whereby the core domain rocks up and down to allow the ion to pass through, after being very sensitively bound and verified. This rocking is not driven by ATP or other outside power, but just by brownian motion, as gated by the ion binding and unbinding steps.
 |
| Drilling a little closer into the target ion binding site of SLC26A6. On right is shown Cl- in green, center, with a few of the amino acids that coordinate its specific, but transient, binding in the core domain pocket. |
They draw contrasts between these very closely related channels, in that the binding pocket is deeper and narrower in SLC26A9, allowing the smaller Cl- to bind while not allowing HCO3- to bind as well. There are also numerous differences in the structure of the core protein around the channel that they argue allow coupling of HCO3- transport (to Cl- transport in the other direction) in SLC26A6, while SLC26A9 is uncoupled. One presumes that the form of the ion site can be subtly altered at each end of the rocking motion, so that the preferred ion is bound at each end of the cycle.
{kind=link}
While all this work is splitting fine hairs, these are hairs presented to us by evolution. It is evolution that duplicated the precursors to these genes, then retained them while each, over time, developed its fine-tuned differences, including different activities and distinct tissue expression. Indeed, the fully competent, bicarbonate exchanging, SLC26A6 is far more widely expressed, suggesting that SLC26A9 has a more specialized role in the body. To reiterate a point made many times before- having the whole human genome sequenced, or even having atomic structures of all of its encoded proteins, is merely the beginning to understanding what these molecular machines do, and how our bodies really work.
Anne Applebaum
ReplyDeleteGood God!!
It's people like her that are starting WWIII!