As previously noted, the ribosome, which translates our DNA code from mRNA copies into the final protein, is a sort of time capsule from the very distant past in the history of life. When RNA was the dominant molecule and proteins and DNA were just getting a foothold, or in the case of DNA, may not have been invented yet at all. It is a bit like, if in the history of civilization, metalworking was stuck at the blacksmithing stage. To make up for the lack of efficiency while all other areas of the economy zoomed ahead into the industrial age (and remained completely dependent on the smith's work) they could only increase production by vast increases in scale, making blacksmithing the predominant industry of the whole economy. In bacteria, ribosomes make up over a quarter of dry cell mass.
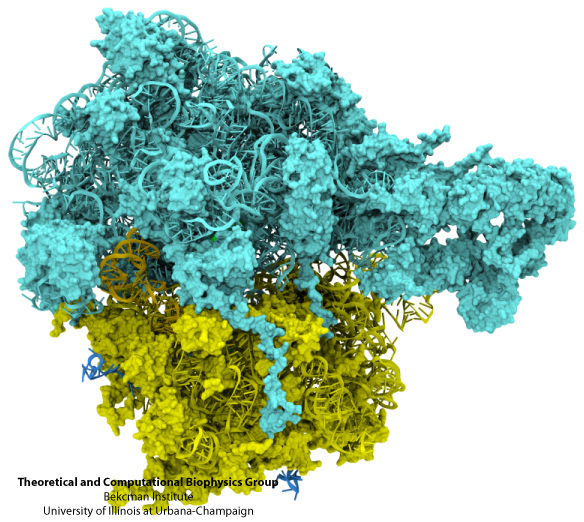 |
| The ribosome, with RNA looking like train tracks and proteins like bits of foam. The large subunit is in aqua, and the small subunit in yellow. |
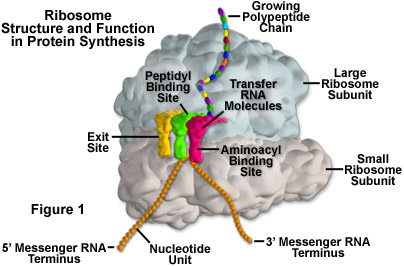
The ribosome is at its core an RNA enzyme, not a protein enzyme. In later eras it became festooned with proteins around its outside- there are over 80 of them in eukaryotes, but they make up only 40% of its weight and have only incidental roles. The core catalytic actions of the ribosome remain RNA-based, taking tRNAs charged with amino acids, (the "L"-shaped molecules in fluorescent colors, below), matching them to mRNA codons as they get threaded through the machine, and linking their amino acids into a growing protein chain. It is a triple RNA nexus, just as one would imagine originally happened if RNA came first in the history of life, and proteins came second, with DNA a distant third.
 |
Why did RNA come first? Because it is a molecule that combines the essential elements of biology- the ability to adopt moderately stable shapes and do things like catalyze reactions, and the ability to be copied, be mutated, and evolve. Unfortunately, it is terrible at all of these roles, but in the land of the blind, the one-eyed molecule is king. So over time, better molecules came along to usurp both of the key roles of biology: DNA to store genetic information in a more stable way, and proteins to form structures and catalysts with far, far greater facility than RNA.
But obviously, the transition from one to the other was tortured and complex, and in the case of protein synthesis, it never really happened. We are left with the hulk that is the ribosome, an enormous and inefficient relic. But that is not all. A paper recently delved into the process that produces ribosomes themselves, which is another story of waste and inefficiency. They found that some of its consituents are related to tRNA processing, showing unexpected relations among the actors in this ancient process. The findings are pretty minor, but it is a nice opportunity to think about this odd corner of biology.
Ribosome production is so onerous that it typically occupies its own cellular zone- the nucleolus, a sub-compartment of the nucleus in eukaryotes. This is where the DNA genes for the ribosomal RNA (typically present in the DNA in many copies, to better ramp up production) congregate and get transcribed by their very own RNA polymerase. This is also where many processing steps happen with the participation of proteins and RNAs imported from outside the nucleolus, like chopping up the full-length RNA into a few pieces, chemical modifications of certain RNA positions, addition of proteins and other RNAs, and the unusual chemical modifications of those proteins in various places. Much of this processing is guided by yet other RNAs that are complementary to various portions of the ribosomal RNA, and have to be pried back off the structure with helicases later on (see below). And then in the end, the whole mess is transported out of the nucleus through nuclear pores that are barely large enough to accommodate it.
I should add that research on ribosomal processing is something of a backwater in molecular biology. The complexity is daunting and technically difficult to deal with, and the whole process is sort of "housekeeping" for the cell, not involved in cancer, development, cognition, or other exciting issues. But still, the advancing tools of the field allow progress on all fronts. Here, the researchers solve the atomic structure of a complex of two proteins, Rrp7, and a partner, Utp22, that seem to be two of those transiently acting proteins that help the ribosome along to maturity- part of a vast support staff.
 |
| Structure of the Utp22+Rrp7 complex, with Rrp7 on the right side in brown and purple. This is the side that binds to the developing ribosome. The D# modules are all part of Utp22. Each partner of this complex is essential for its further ribosomal maturation. |
Rrp7 contains the key region that binds the small complex to an RNA site within what later becomes the small subunit of ribosomal RNA, but it requires a guide RNA (snR30) which has to bind the rRNA and stabilize a particular kink before it can do so. What happens next? That part is less clear. Utp22 may bring along other proteins and enzymes, but it doesn't have enzymatic activity itself. The authors do not delve into this aspect, unfortunately. Its role may actually not be to do anything in particular for the ribosome, but simply to bind this intermediate state, and then when released to regulate a set of ribosomal protein genes in a system that makes their gene transcription responsive to the overall level of ribosome processing and production.
"The complex structure of Utp22 and Rrp7 shows that they are unlikely to possess any enzymatic activity and that they rather function as an essential building block in the 90S preribosome."
 |
| A small area of the ribosomal RNA, (18S), showing a few sites where Rrp7 binds and can be cross linked (green nucleotides), and also showing the guide RNA, snR30 in blue, which transiently binds to and stabilizes the ribosomal RNA, and is essential for subsequent Rrp7 binding. |
What the authors find interesting is the evolutionary history of Utp22, which is that it developed from a tRNA processing enzyme called the CCA-adding enzyme. This enzyme adds a special three-nucleotide end to all tRNAs which is not originally present in their DNA code. Why? Who knows- probably another historical hack that came along prior to the rise of what we regard as more orderly & conventional (dogmatic, one might even say!) molecular biology. A copy of this enzyme was re-purposed to become this small subunit ribosomal RNA processing factor, eventually losing both its catalytic activity (of adding nucleotides to RNA ends) and its RNA-binding function (which was taken over by Rrp7).
This is only a taste of the intricacies of this field. Biology is full of hacks and workarounds, also called adaptations, but the ribosomal system takes the cake for its overall cost, its focus on RNA as the central and primordial molecule, and its byzantine chemical and macromolecular complexity. Life is certainly weird tech, but it isn't always high tech.
- Earliest life pushed back by about 70 million years.
- We need a much bigger discussion of where our economy is going.
- What ever happened to public education?
- Krugman talks up MMT.
- Krugman is making at least one person's head explode.
- Whom did QE help?
- More reasons to hate Pakistan, if any are needed.
- The Senate is still an unconstitutional and broken institution.
- What's going on with unions? We need a new labor regime.
- Education is not the cause of the recent rise in economic inequality.
- The endangered species act is 40 years old. Yay!
No comments:
Post a Comment
Thank you for commenting!