Cell nuclei are consistently sized with respect to cell volume, and pleasingly round. How does that happen?
An interesting question in biology is why things are the size they are. Why are cells so small, and what controls their size? Why are the various organelles within them a particular size and shape, and is that controlled in some biologically significant way, or just left to some automatic homeostatic process? An interesting paper come out recently about the size of the nucleus, home of our DNA and all DNA-related transactions like transcription and replication. (Note to reader/pronouncer: "new clee us", not "new cue lus".)
 |
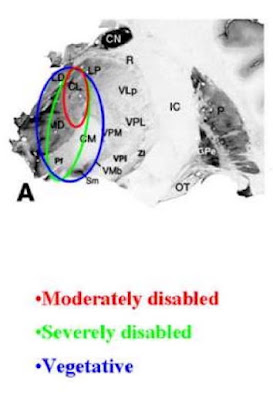
| The nucleus, with parts labeled. Pores are large structures that control traffic in and out. |
The nucleus is surrounded by a double membrane (the nuclear membrane) studded with structurally complex and interesting pores. These pores are totally permeable to small molecules like ions, water, and very small proteins, but restrict the movement of larger proteins and RNAs, and naturally, DNA. To get out, (or in), these molecules need to have special tags, and cooperate with nuclear transport proteins. But very large complexes can be transported in this way, such as just-transcribed RNAs and half-ribosomes that get assembled in the nucleolus, a small sub-compartment within the nucleus (which has no membrane, just a higher concentration of certain molecules, especially the portion of the genomic DNA that encodes ribosomal RNA). So the nuclear pore is restrictive in some ways, but highly permissive in other ways, accommodating transmitted materials of vastly different sizes.
 |
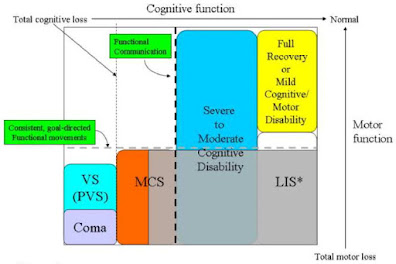
| Nuclear pores are basket-shaped structures that are festooned, particularly inside the channel, with disordered phenylalanine/glycine rich protein strands that act as size, tag, and composition-based filters over what gets through. |
The channels of nuclear pores have a peculiar composition, containing waving strands of protein with repetitive glycine/phenylalanine composition, plus interspersed charged segments (FG domains). This unstructured material forms a unique phase, neither oily nor watery, that restricts the passage of immiscible molecules, (i.e., most larger molecules), unless accompanied by partners that bind specifically to these FG strands, and thus melt right through the barrier. This mechanism explains how one channel can, at the same time block all sorts of small to medium sized RNAs and proteins, but let through huge ribosomal components and specifically tagged and spliced mRNAs intended for translation.
But getting back to the overall shape and size of the nucleus, a recent paper made the case in some detail that colloid pressure is all that is required. As noted above, all small molecules equilibrate easily across the nuclear membrane, while larger molecules do not. It is these larger molecules that are proposed to provide a special form of osmotic pressure, called colloid osmotic pressure, which gently inflates the nucleus, against the opposing force of the nuclear membrane's surface tension. No special mechanical receptors are needed, or signaling pathways, or stress responses.
The paper, and an important antecedent paper, make some interesting points. First is that DNA takes up very little of the nuclear volume. Despite being a huge molecule (lengthwise), DNA makes up less than 1% of nuclear volume in typical mammalian cells. Ribosomal RNA, partially constructed ribosomal components, tRNAs, and other materials are far more abundant and make up the bulk of large molecules. This means that nuclear size is not very sensitive to genome copy number, or ploidy in polyploid species. Secondly, they mention that a vanishingly small number of mutants have been found that affect nuclear size specifically. This is what one would expect for a simple- even chemical- homeostatic process, not dependent on the usual signaling pathways of cellular stress, growth regulation, etc., of which there are many.
Where does colloid osmotic pressure come from? That is a bit obscure, but this Wiki site gives a decent explanation. When large molecules exist in solution, they exclude smaller molecules from their immediate vicinity, just by taking up space, including a surface zone of exclusion, a bit like national territorial waters. That means that the effective volume available to the small solutes (which generally control osmotic pressure) is slightly reduced. But when two large molecules collide by random diffusion, the points where they touch represent overlapping exclusion zones, which means that globally, the net exclusion zone from large molecules has decreased, giving small solutes slightly more room to move around. And this increased entropy of the smaller solutes drives the colloid osmotic pressure, which rises quite rapidly as the concentration of large molecules increases. The prior paper argues that overall, cells have quite low colloid osmotic pressure, despite their high concentrions of complex large molecules. They are, in chemical terms, dilute. This helps our biochemistry do its thing with unexpectedly rapid diffusion, and is explained by the fact that much of our molecular machinery is bound up in large complexes that reduce the number of independent colloidal particles, even while increasing their individual size.
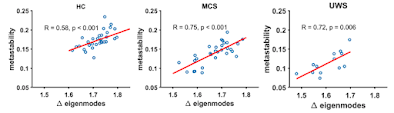
So much for theory- what about the experiment? The authors used yeast cells (Schizosaccharomyces pombe), which are a common model system. But they have cell walls, which the researchers digested off before treating them with a variety of osmolytes, mostly sorbitol, to alter their osmotic environment (not to mention adding fluorescent markers for the nuclear and plasma membranes, so they could see what was going on). Isotonic concentration was about 0.4 Molar (M) sorbitol, with treatments going up to 4M sorbitol (hypertonic). The question was.. is the nucleus (and the cell as a whole) a simple osmometer, reacting as physical chemistry would expect to variations in osmotic pressure from outside? Recall that high concentrations of any chemical outside a cell will draw water out of it, to equalize the overall water / osmotic pressure on both sides of the membrane.
 |
| Schizosaccharomyces pombe are oblong cells (left) with plasma membrane marked with a green fluorescent marker, and the nuclear membrane marked with a purple fluorescent marker. If one removes the chitin-rich cell wall, the cells turn round, and one can experiment on their size response to osmotic pressure/treatment. Hypertonic (high-sorbitol, top) treatment causes the cell to shrink, and causes the nucleus to shrink in strictly proportional fashion, indicating that both have simple composition-based responses to osmotic variation. |
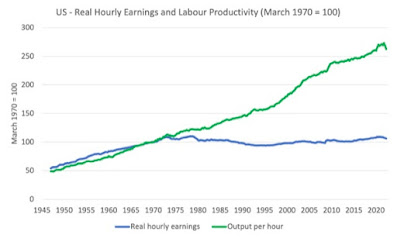
They found that not only does the outer cell membrane shrink as the cell comes under hypertonic shock, but the nucleus shinks proportionately. A number of other experiments followed, all consistent with the same model. One of the more interesting was treatment with leptomycin B (LMB), which is a nuclear export inhibitor. Some materials build up inside the nucleus, and one would expect that, under this simple model of nuclear volume homeostasis, the nuclei would gradually gain size relative to the surrounding cell, breaking the general observation of strict proportionality of nuclear to cell volumes.
 |
| Schizosaccharomyces pombe cells treated with a drug that inhibits nuclear export of certain proteins causes the nuclear volume to blow up a little bit, relative to the rest of the cell. |
That is indeed what is seen, not really immediately discernable, but after measuring the volumes from micrographs, evident on the accompanying graph (panel C). So this looks like a solid model of nuclear size control, elegantly explaining a small problem in basic cell biology. While there is plenty of regulation occuring over traffic into and out of the nucleus, that has critical effects on gene expression, translation, replication, division, and other processes, the nucleus can leave its size and shape to simple biophysics and not worry about piling on yet more ornate mechanisms.
- About implementing the climate bill and related policies.
- We should have given Ukraine to Russia, apparently. Or something.
- Big surprise- bees suffer from insecticides.